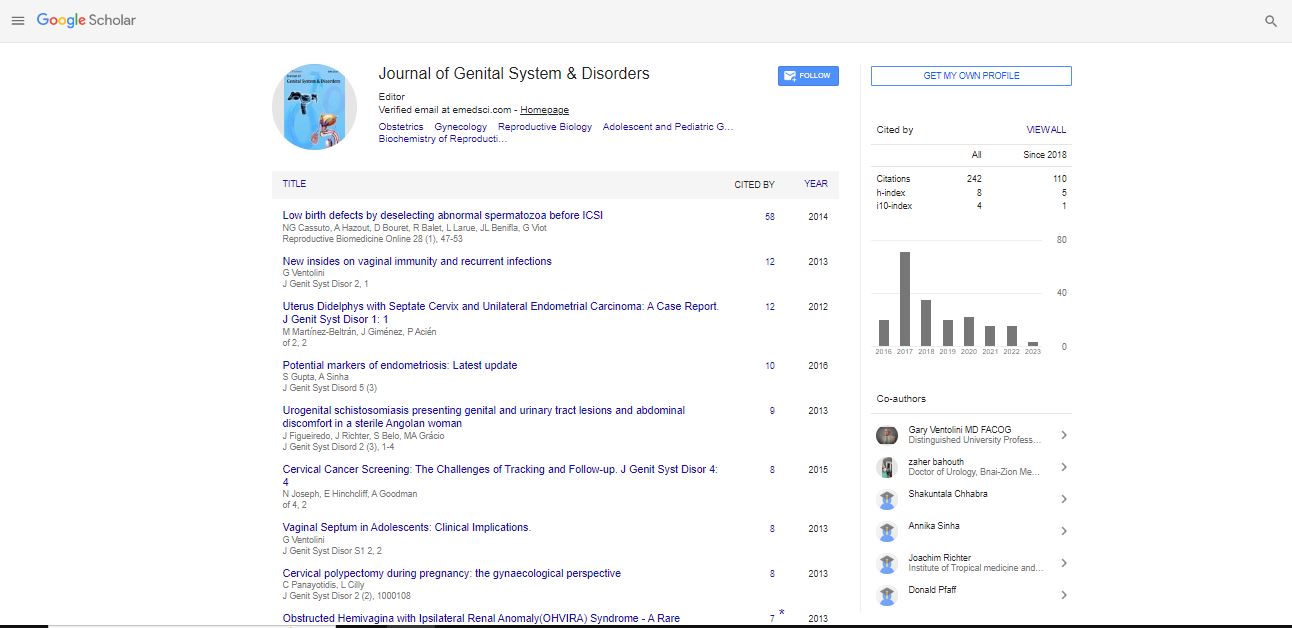
Spanish
Spanish  Chinese
Chinese  Russian
Russian  German
German  French
French  Japanese
Japanese  Portuguese
Portuguese  Hindi
Hindi Research Article, J Genit Syst Disor Vol: 5 Issue: 1
Characterization of Toll-Like Receptor 9 expression during Mouse Preimplantation Development
| Sasaki K1, Tanaka A1, Nagatomo H1, Ogawa H2, Kobayashi K3,Takahashi M1 and Kawahara M1* | |
| 1Laboratory of Animal Breeding and Reproduction, Graduate School of Agriculture, Hokkaido University, Sapporo 060-8589, Japan | |
| 2Department of BioScience, Tokyo University of Agriculture, 1-1-1 Sakuragaoka, Setagaya-ku, Tokyo 156-8502, Japan | |
| 3Laboratory of Dairy Food Science, Research Faculty of Agriculture, Hokkaido University, Sapporo 060-8589, Japan | |
| Corresponding author : Manabu K, PhD Laboratory of Animal Breeding and Reproduction, Graduate School of Agriculture, Hokkaido University, Sapporo 060-8589, Japan Tel: +81-11-706-2541 E-mail:khara@anim.agr.hokudai.ac.jp |
|
| Received: December 17, 2015 Accepted: January 08, 2016 Published: January 15, 2016 | |
| Citation: Sasaki K, Tanaka A, Nagatomo H, Ogawa H, Kobayashi K, et al. (2016) Characterization of Toll-Like Receptor 9 expression during Mouse Preimplantation Development. J Genit Syst Disor 5:1. doi:10.4172/2325-9728.1000145 |
Abstract
Toll-like receptor 9 (TLR9) is a pathogenic sensing receptor that binds to various pathogenic components, recognizing foreign genomic DNA. Numerous studies have reported the functions of TLR9 in somatic cells, but its feature in germ cells and early embryos has not been well understood. In this study, to characterize TLR9 during preimplantation development, we investigated the mRNA and protein expression dynamics of TLR9 in mouse oocytes and embryos.
Keywords: TLR9; Expression; Oocyte; Preimplantation embryo; Trophectoderm
Keywords |
|
| TLR9; Expression; Oocyte; Preimplantation embryo; Trophectoderm | |
Abbreviations |
|
| ESC: Embryonic Stem Cell; ICM: Inner Cell Mass; IFN: Interferon; PRR: Pattern Recognition Receptor; TLR: Toll-Like Receptor; TE: Trophectoderm; TSC: Trophoblast Stem Cell | |
Introduction |
|
| In mammals, the innate immune system plays essential roles in maintaining homeostasis through numerous pathways involving Tolllike receptor (TLR) signaling [1]. Pathogen recognition occurs during the first step of innate immunity, which is triggered by the sensing of pattern recognition receptors (PRRs) derived from pathogens. The TLR molecules are flagship members of this system and counteract pathogenic infections in host animals. In mice, the TLR family consists of 12 members (TLR1–9 and TLR11–13). Each member recognizes specific molecular components derived from pathogens [2-14]. After pathogen recognition, TLRs stimulate downstream signaling pathways to induce type I interferon-α/β (IFN-α/β) and inflammatory cytokine secretion [1]. During mammalian post implantation development, TLRs are expressed in both embryonic and placental tissues in mid and late pregnancy [15,16]. | |
| In preimplantation development, TLR members were expressed in the cell surfaces in embryos from the 2-cell to blastocyst stages, of which TLR2 and TLR4 were localized mainly in the trophectoderm (TE) of blastocyst [17]. The first differentiation occurs at the blastocyst stage into 2 types of cell lineage: inner cell mass (ICM) and TE. The former mainly generates embryo proper; on the other hand, the latter gives rise to extra embryonic tissue such as placenta. In addition, TE cell-derived trophoblast stem cells (TSCs) expressed Tlr1–6 mRNA but not Tlr9, and also expressed antiviral genes including IFN-β by TLR3 agonist stimuli [18]. This indicates that TE and TSCs are potent to show antipathogenic responses. On the other hand, certain types of TLRs expressed in embryonic stem cells (ESCs) could not activate downstream signaling pathways to IFN-α/β [19]. Therefore, TLRs appear to show cell type-specific characteristics in ESCs and TSCs. However, there is not enough knowledge about pathogen sensor TLR9 in oocytes and early embryos in mammals. | |
| We here focused on the characteristics of TLR9 in mouse oocytes and preimplantation embryo. We first verified Tlr9 mRNA and TLR9 protein expression in mouse preimplantation embryos from the zygotic stage to the blastocyst stage. Furthermore, we observed the localization pattern of the TLR9 protein by immunofluorescence analysis. The localization of the TLR9 protein showed the TE-predominant pattern at the blastocyst stage. | |
| Consequently, we used ESCs and TSCs to determine the TLR9 mRNA expression in a lineage-specific manner. Our results demonstrated the expression of Tlr9 transcripts/TLR9 proteins during preimplantation development, and further ensured the TE lineage-predominant expression at the blastocyst stage. | |
Materials and Methods |
|
| Mice | |
| We collected cumulus-oocyte complexes (COCs) and spermatozoa from 10– 14-week-old C57BL/6N mice. For super ovulation treatment, female mice were administrated 7.5 IU equine chorionic gonadotropin (eCG; ASKA Pharmaceutical, Tokyo, Japan). Forty-eight hours after administration, 7.5 IU human chorionic gonadotropin (hCG; ASKA Pharmaceutical) was injected into the mice. All experimental procedures were approved by the Animal Care and Use Committee of Hokkaido University | |
| In Vitro fertilization and embryo culture | |
| Spermatozoa were collected from the caudal epididymis of adult male C57BL/6N mice and cultured in human tubular fluid (HTF) medium for 1.5 hr [20,21]. COCs were collected from the ampulla of female oviducts into HTF medium at 16 hr post-hCG injection and inseminated with incubated spermatozoa for 6 hr in HTF medium at 37°C in 6% CO2. After insemination, fertilized oocytes were washed in M2 medium to remove cumulus cells and spermatozoa and subsequently cultured in M16 medium [22,23]. | |
| Preparation of oocyte and embryo | |
| Oocytes and preimplantation embryos were prepared at each developmental stage for quantitative PCR and immunofluorescence. Oocytes in the metaphase stage of the second meiosis (Oo) were obtained from super ovulation-treated female mice. Additionally, 1-cell (Zy), 2-cell (2C), 4-cell (4C), 8-cell (8C), morula (M), and blastocyst (B) stage embryos were collected at 6, 30, 48, 60, 72, and 84 h post-fertilization, respectively. The zona pellucida of all oocytes and embryos were removed by using acidic Tyrode’s solution for 10–20 sec at 20°C. | |
| Cell culture | |
| ESCs were derived from C57BL/6J strain blastocysts (XY) and cultured in 2i/LIF media [24]. TSCs were established from B6D2F1 (C57BL/6 × DBA/2) cultured in TS medium TS medium [25] (RPMI 1640 (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (JRH Biosciences Inc.), 1 mM sodium pyruvate (Thermo Fisher Scientific), 100 μM β-mercaptoethanol (Sigma), 2 mM l-glutamine (Thermo Fisher Scientific), 100 U/ml penicillin and 100 μg/ml streptomycin (Thermo Fisher Scientific)), containing 25 ng/mL fibroblast growth factor 4 (FGF4; PeproTech EC, Rocky Hill, NJ, USA) and 1 mg/mL heparin (Sigma-Aldrich, St. Louis, MO, USA). TSCs were differentiated by removing FGF4, heparin, and feeder cells, and were collected at 2, 4, and 6 days postdifferentiation (D2, D4, and D6, respectively). The cultivation and differentiation of TSCs were conducted as described previously [26]. | |
| RNA isolation and reverse-transcription | |
| Total RNAs from oocytes and embryos were extracted by using the ReliaPrep RNA Cell Miniprep System (Promega, Madison, WI, USA) and reverse-transcribed into cDNAs by using ReverTra Ace qPCR RT Master Mix (Toyobo, Osaka, Japan). The number of oocytes and embryos for RNA extraction were shown as follows; oocyte: 50, zygote: 50, 2-cell: 50, 4-cell: 30, 8-cell: 30, morula: 20, and blastocyst: 20. Next, cDNA concentrations were standardized to 1 μg/μL by using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific). For ESCs and TSCs, total RNA was isolated by using ISOGEN II (Nippon Gene, Tokyo, Japan) and digested by RQ1 RNase-free DNase (Promega). ESCs and TSCs used for RNA extraction were 105 and 4 × 104 cells, respectively. First-strand cDNA was synthesized from 1 μg of total RNA by using SuperScript III (Life Technologies, Carlsbad, CA, USA) using the oligo dT primer. | |
| RT-PCR | |
| Quantitative RT-PCR was performed by using a LightCycler 480 (Roche Diagnostics, Basel, Switzerland) using standardized cDNA. Three independent experiments were replicated for each developmental stage. The reactions were performed by using THUNDERBIRD SYBR qPCR Mix (Toyobo). Annealing temperature was 60°C in all reactions. Relative expression levels were calculated relative to that of the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (Gapdh) in all samples. We used primer pairs (5’– ATGGTTCTCCGTCGAAGGACT–3’ and 5’–CAGGTGGTGGATACGGTTGG–3’; 234 base pairs) to amplify Tlr9 (GenBank ID: NM_031178), and another primer set (5’– GTCGTGGAGTCTACTGGTGTC–3’ and 5’– GAGCCCTTCCACAATGCCAAA–3’; 240 base pairs) for GAPDH (GenBank ID: NM_008084) [27]. Alternatively, 102–108 copies of PCR amplicons, purified by using the Wizard SV Gel and PCR Clean-Up System (Promega), were subjected to quantitative RT-PCR analysis as external standard samples. Tlr9 and Gapdh expressions were also evaluated in ESCs, undifferentiated TSCs, and differentiated TSCs at D2–D6, by RT-PCR by using TaKaRa Ex Taq (Takara, Shiga, Japan). The cycling profile for Tlr9 comprised an initial denaturation step for 94°C for 5 min, followed by 40 cycles at 94°C for 30 sec, 60°C for 30 sec, 72°C for 30 sec, and a final extension step at 72°C for 5 min. For the detection of house-keeping Gapdh, PCR was performed by 35 cycles. The PCR amplicons were electrophoresed in 2% agarose gel and stained with ethidium bromide to visualize the expression of Tlr9 transcripts in cultured cells. Three independent experiments were replicated. | |
| Western blotting | |
| To assess TLR9 protein expression, 50 oocytes and blastocysts dissolved in sample buffer were electrophoresed by SDS-PAGE in an 8% polyacrylamide gel and then transferred onto polyvinylidene fluoride membranes. The liver tissue retrieved from adult mouse after fine mincing was prepared by the same procedure in oocytes and embryos, followed by SDS-PAGE. The membranes were treated with blocking buffer (4% skim milk and 0.2% Tween 20) for 1 hr, and then reacted with monoclonal TLR9 antibody (NBP2-24729; 1:125; Novus Biologicals, Littleton, CO, USA) at 4°C overnight. The membrane was washed with PBS containing 0.05% Tween 20 and treated with anti-mouse IgG-peroxidase antibody from goat (A0168; 1:2500; Sigma-Aldrich). Next, TLR9-chemiluminescence was detected by the Luminata Forte Western HRP substrate (Millipore, Billerica, MA, USA) by using a Bio-Rad ChemiDoc XRS+ (Bio-Rad, Hercules, CA, USA). Expression of β-actin was also detected by using monoclonal anti-β-actin (Clone AC-15; Sigma-Aldrich) as an internal control. | |
| Immunofluorescence | |
| Oocytes and preimplantation embryos were fixed in 4% (w/v) paraformaldehyde (Wako, Osaka, Japan) for 1 hr at room temperature and then permeabilized by 0.2% (w/v) Triton X-100 (Sigma-Aldrich) for 1 hr at room temperature. Subsequently, the samples were treated with blocking solution (3.0% [w/v] bovine serum albumin [BSA; Sigma-Aldrich] in PBS) for 1 hr at room temperature, and then incubated with TLR9 antibody (1:200) diluted in blocking solution at 4°C overnight. After washing 3 times with washing solution (0.1% [v/v] Triton X-100, 0.3% [w/v] BSA in PBS), the samples were incubated with Alexa Fluor 488 goat anti-mouse IgG (H+L) (A- 11001; 1:300; Life Technologies) diluted in blocking solution for 1 hr at room temperature. Subsequently, the samples were washed 5 times with washing solution and then mounted by using VECTASHIELD Mounting Medium with DAPI (H-1200; Vector Laboratories, Inc., Burlingame, CA, USA). Fluorescent signals were visualized using the EVOS® Cell Imaging System (Advanced Microscopy Group, Mill Creek, WA, USA), and confocal laser scanning microscopy with a Leica TCS SP5 II (Leica Camera, Solms, Germany). | |
| Dissection of blastocysts | |
| To precisely determine the localization of TLR9 protein, blastocysts were dissected mechanically into an inner cell mass (ICM) side and trophectoderm (TE) side by using a micromanipulator equipped with a microsurgical blade (FEATHER, Osaka, Japan). Cells on the ICM side contained ICM and parts of TE cells, which adhered closely to the ICM at the mural TE. The TE side contained only TE cells surrounding the inner blastocoel, known as the polar TE. Each sample was fixed and subjected to immunofluorescence analysis. | |
Statistical Analysis |
|
| Statistical analysis of the quantitative PCR results for comparison was carried out by using the Fisher protected least significant difference test by using STATVIEW (Abacus Concepts, Inc., Berkeley, CA, USA). Experimental data are presented as the mean ± standard errors. P values of < 0.05 were considered statistically significant. | |
Results |
|
| Expression profiles of TLR9 during preimplantation development | |
| We first examined the Tlr9 mRNA expression in mouse oocytes and preimplantation embryos at each developmental stage (Figure 1A). The expression levels of Tlr9 mRNA were higher in the oocytes and embryos at the 1-cell stage, whereas expression rapidly decreased during development. We did not detect the Tlr9 mRNA expression at the morula stage but observed low expression at the blastocyst stage. To further investigate the TLR9 protein expression, we performed western blotting analysis for oocyte, blastocyst, and liver tissue as positive control [28]. Expectedly, we detected the TLR9 protein expression in both oocyte and blastocyst (Figure 1B). Next, we investigated the intracellular localization of TLR9 protein by immunofluorescence analysis (Figure 2). The TLR9 protein was detected in both the cytoplasm and nucleus in oocytes and preimplantation embryos during all developmental stages, although the fluorescent intensity in nucleus was slightly weaker. Furthermore, the fluorescent signals of TLR9 mostly disappeared at the 4-cell stage. Interestingly, at the blastocyst stage, TLR9 was localized predominantly in the TE (Figure 2). | |
| Figure 1: Expression profiling of Tlr9 mRNA and TLR9 protein during preimplantation Development. A] qRT-PCR analysis of Tlr9 transcripts in oocytes and preimplantation embryos at each developmental stage. Relative expression was calculated relative to the expression level in an oocyte. Error bar indicates standard error of the mean. B] Western blotting of TLR9 in oocytes and blastocysts. As a positive control, the same experiment was conducted in the liver. β-Actin was analyzed as an internal control. Oo, oocyte; Zy, 1-cell embryo; 2C, 2-cell embryo; 4C, 4-cell embryo; 8C, 8-cell embryo; M, morula embryo; B, blastocyst embryo; Li, liver; ND, not detected. | |
| Figure 2: Intracellular localization of TLR9 during preimplantation development. Immunofluorescence analysis in oocytes and preimplantation embryos. TLR9 (green) was labeled with Alexa Fluor® 488, and the nucleus was stained with DAPI (blue). Scale bar = 100 µm. As a negative control, oocytes were examined without the primary antibody (negative). Oo, oocyte; Zy, 1-cell embryo; 2C, 2-cell embryo; 4C, 4-cell embryo; 8C, 8-cell embryo; M, morula embryo; B, blastocyst embryo. | |
| Characterization of tlr9 localizations at the pronuclear and blastocyst stages | |
| Based on the results shown in Figure 2, we precisely investigated the localization of TLR9 by confocal microscopic analysis at both the pronuclear and blastocyst stages. At the pronuclear stage, there were no differences in the localization pattern of TLR9 between male and female pronuclei-surrounding areas (Figure 3A). We next analyzed the space-predominant localization of TLR9 in the whole blastocyst and dissected blastocysts, including the ICM and TE sides of the blastocyst. In the whole blastocyst, TLR9 expression was biased to the opposite side of the ICM (Figure 3B). In dissected blastocysts, TLR9 expression was observed in the TE side, whereas expression was moderate in the ICM side (Figure 3B). This indicated the TEpredominant expression of TLR9 during the blastocyst stage. | |
| Figure 3: Locational characterization of TLR9 in zygote and blastocyst embryos. [A] Confocal images of TLR9 (green) and pronuclei (blue) in zygote at 6 h post-fertilization. Scale bar = 20 µm. [B] Cell lineage-dependent TLR9 characteristics in the blastocyst embryos. Blastocyst embryo was divided into ICM and TE sides with microblade. Whole indicates non-dissected embryo. Each sample was subjected to immunofluorescence analysis of TLR9 (green) and nucleus (blue). Scale bar = 50 µm. | |
| Gene Expression of Tlr9 mRNA in TSCs | |
| In order to definitely determine TE-predominant expression of TLR9, we analyzed the expression of Tlr9 mRNA in TSCs and ESCs (Figure 4). TSCs maintain their multipotency to all types of cells for placentation with the supplement of FGF4 and heparin into the culture medium [29]. We previously confirmed that the TS cell line used here displayed the trophoblast giant cell-like gene expression profile at 6 days post differentiation (D6) [26]. The Tlr9 mRNA expression was detected in undifferentiated TSCs (D0) but not in ESCs (Figure 4). Furthermore, the Tlr9 mRNA expression was confirmed on D2, D4, and D6 without reference to the progress of TSC differentiation. These results supported the trophoblast cell lineage-predominant expression of Tlr9 mRNA. | |
| Figure 4: Tlr9 expressions in different cell lineages. RT-PCR analysis of Tlr9 was demonstrated in 5 cell types: embryonic stem cells (ESCs), undifferentiated trophoblast stem cells (TSCs D0), and differentiated TSCs at 2, 4, and 6 days post-differentiation (TSCs D2, D4, and D6). No Tlr9 expression was detected in ESCs. Gapdh expression was assessed as an internal control. | |
Discussion |
|
| We here addressed TLR9 mRNA expression and protein localization patterns during mouse preimplantation development. qRT-PCR analysis revealed that Tlr9 mRNA was expressed in oocytes and preimplantation embryos, except for the morula stage. The expression level decreased from the 2-cell to morula stages, indicating that the Tlr9 mRNA expression before the morula stage was obviously derived from oocyte. Tlr9 rapidly declined post-fertilization, differing from the result of the TLR9 protein localization by immunostaining. TLR9 proteins were present during all the developmental stages. This TLR9 expression pattern might be due to the re-expression of its mRNA at the blastocyst stage. TLR9 stimulates anti-pathogenic signaling by sensing unmethylated pathogenic DNA [7]. However, TLR9 was localized to the cytoplasm of each blastomere and not inside the pronucleus (Figure 3A). In somatic cells such as dendritic cells and macrophages, it is known that TLR9 molecules are localized in the endosome to be kept away from genomic DNA and not to recognize it [30]. Therefore, our results indicated that TLR9 within ooplasm did not contribute to the selective recognition of “exogenous” paternal DNA during the fertilization. | |
| We next focused on the remarkable changes of TLR9 localization observed at the blastocyst stage when the first differentiation into ICM and TE occur (Figure 3B). The TE-predominant expression of Tlr9 was also observed in TSCs but not in ESCs (Figure 4). On the other hand, the previous study showed that Tlr9 mRNA was expressed in neither ESCs nor TSCs [18]. This contradiction might be due to the different cell lines used for experiments and/or the low level of TLR9 mRNA/protein expression in the blastocyst (Figure 1). The copy number of Tlr9 transcripts in both blastocysts and TSCs was extremely low, which might make it difficult to detect the Tlr9 expression. ESCs are sensitive to infections by RNA viruses, as these cells do not secrete IFN and inflammatory cytokines [19]. In contrast, trophoblasts possess antiviral responses via TLRs and protect pathogen-sensitive ICM cells from pathogenic infection [17,18]. Our observations regarding TLR9 were consistent with their reports. | |
| The TE-predominant TLR9 expression maintained in differentiated TSCs after the removal of both FGF4 and heparin, indicating that TLR9 might contribute to the antiviral function at post-implantation stage in trophoblast-derived cell lineages. Although further experiments are required to determine the biological significance of TLRs, TLR9 is definitely present within both oocytes and embryos, and its expression is mainly localized to the trophectoderm as well as the other TLRs. Our findings would provide basis for further understanding the molecular mechanisms underlying a diversity of TLRs during mouse preimplantation development. | |
Acknowledgments |
|
| We thank Prof. Fumio Nakamura (Laboratory of Animal By-Product Science, Research Faculty of Agriculture, Hokkaido University, Sapporo, Japan) for providing critical advice regarding immunofluorescence analysis and western blotting analysis. Part of this work was supported by Grant-in-Aid for Research on Priority Area to Young Scientists (B) to M. K. (No. 24780265) and Grant-in-Aid for Scientific Research (C) to M. T. (No. 24580439) from the Ministry of Education, Culture, Sports, Science and Technology of Japan. | |
References |
|
|
|