Spanish
Spanish  Chinese
Chinese  Russian
Russian  German
German  French
French  Japanese
Japanese  Portuguese
Portuguese  Hindi
Hindi Research Article, J Nanomater Mol Nanotechnol Vol: 5 Issue: 4
Macrophage Cells Secrete Specific Cytokines and Accumulate Activated Interferon Regulatory Factor 3 after Multi-Walled Carbon Nanotube Exposure
| Kevin D Houston1*, Nathan H Mack2, Stephen K Doorn3 and Min S Park4 | |
| 1Department of Chemistry and Biochemistry, New Mexico State University, Las 5 Cruces, NM 88003, USA | |
| 2Chemistry Division, Los Alamos National Laboratory, Los Alamos, NM 87545, USA | |
| 3Center for Integrated Nanotechnologies, Los Alamos National Laboratory, Los 8 Alamos, NM 87545, USA | |
| 4Center for Microalgal Biotechnology and Biofuels Institute of Hydrobiology, Chinese Academy of Sciences Wuhan, China | |
| Corresponding author : Kevin D Houston Department of Chemistry and Biochemistry, New Mexico State University, Las 5 Cruces, NM 88003, USA E-mail: khouston@nmsu.edu |
|
| Received: May 19, 2016 Accepted: August 08, 2016 Published: August 14, 2016 | |
| Citation: Houston KD, Mack NH, Doorn SK, Park MS. (2016) Macrophage Cells Secrete Specific Cytokines and Accumulate Activated Interferon Regulatory Factor 3 after Multi-Walled Carbon Nanotube Exposure. J Nanomater Mol Nanotechnol 5:4. doi:10.4172/2324-8777.1000195 |
Abstract
Macrophage Cells Secrete Specific Cytokines and Accumulate Activated Interferon Regulatory Factor 3 after Multi-Walled Carbon Nanotube Exposure
The health consequences of human exposure to carbon-based nanomaterials are not fully understood and the effects that such materials have on the immune system have not been adequately characterized. To determine if the innate immune system is modulated by exposure to carbon-based nanomaterials, the extracellular accumulation of an array of cytokines was measured in cell culture media obtained from mouse macrophage cells (RAW264.7) following exposure to carbon nanotubes (CNTs) or fullerene (C60). Accumulation of a specific subset of cytokines was observed after exposure to multi-walled CNTs (MWCNT), but was not observed when cells were exposed to single-walled CNTs (SWCNTs) or C60. Additionally, the accumulation of the activated (phosphorylated) form of the interferon regulatory factor 3 (IRF3) transcription factor and associated interferon beta (IFNβ) was observed after MWCNT exposure. These data suggest that IRF3 is a mediator of MWCNTactivated signal transduction pathways in macrophage cells. Furthermore, our data show that MWCNTs induce an innate immune response at subtoxic doses and suggests that MWCNT exposure may result in chronic inflammation and compromised immunity.
Keywords: Fullerene; Carbon nanotube; Innate immunity; Cytokine; IRF3
Keywords |
|
| Fullerene; Carbon nanotube; Innate immunity; Cytokine; IRF3 | |
Introduction |
|
| The potential for increased human exposure to engineered nanomaterials is unavoidable owing to ongoing efforts aimed at identifying novel uses for such materials that range from biomedical imaging to solar energy capture [1-4]. Carbon nanotubes (CNTs) have unique properties that make them ideal for biomedical applications such as drug delivery, tissue engineering scaffolds, and for diagnostic biomedical imaging [5-10]. While applications utilizing CNTs hold great promise, potential human health impacts associated with exposing humans to CNTs are poorly understood. | |
| A potential human health impact resulting from CNT use in biomedicine is cytotoxicity. Many research groups have evaluated the cytotoxic potential of CNTs using various model systems and biological endpoints [11]. Currently, it is not clear if single-walled or multi-walled carbon nanotubes (SWCNTs and MWCNTs, respectively) are cytotoxic. Some reports provide evidence that MWCNTs are toxic to certain cell lines while other groups report that MWCNTs are nontoxic [12-16]. Still other reports indicate that the potential for MWCNT cytotoxicity is dependent on residual catalyst or the size of the nanotube with regard to length [17,18]. To further complicate the issue, the MTT assay commonly used to evaluate cytotoxicity was later proven ineffective due to unforeseen chemical interactions between the carbon nanotubes and the reporter precipitate [19]. Also, the cytotoxic effect may be a function of the way the treatment was prepared whether it be sonication or the use of a surfactant [12]. The current data regarding the cytotoxic potential associated with CNT exposure is not conclusive [20]. | |
| There is also evidence that exposure to CNTs impacts other cellular processes. In human embryonic kidney cells (HEK293) and human fibroblast cells, SWCNTs inhibit cell proliferation via modulating the transcription of cell cycle regulating genes and inducing apoptosis [21]. SWCNTs have also been shown to induce ROS generation in human keratinocytes and to increase the levels of oxidative stress markers in other cell types [13,22-24]. Additionally, granuloma formation in rodent lungs and apparent respiratory toxicity in rainbow trout resulting from SWCNT exposure suggest a potential negative human health impact upon exposure to this material [25-28]. Other reports indicate that SWCNTs do not induce acute cytotoxicity in certain cell types, including RAW264.7 cells, and do not affect embryo development in zebrafish [16,19,25,29]. Furthermore, the observed SWCNT-induced biological responses and/or cytotoxicity may be a function of nanotube purity and attachment of functional groups [17,30]. | |
| Carbonaceous nanoparticles including nanotube aggregates have been in our environment for at least 10,000 years as evidenced by the discovery of such materials in ice cores from Greenland [31]. In the case of carbon-based nanomaterials, it is likely that organisms have evolved in presence of these nanomaterials suggesting that the innate immune system may have evolved to respond to such materials. The innate immune system is evolutionarily conserved from plants to mammals and is the first line of host defense against pathogens. Typically, the innate immune response is activated following the binding of pathogen-associated molecular patterns (PAMPs) by pattern recognition receptors (PRRs) that are expressed by many cell types. Activation of PRRs culminates in the production of various chemokines/cytokines. Production and secretion of chemokines/ cytokines is indispensable for an effective innate immune response; they function to “alert” immune response cells of a pathogenic challenge and often activate or serve as chemoattractants for certain immune cells [32,33]. Due to the diversity of PAMPs that are recognized by innate immune system-activating PRRs, it is possible that PRRs may be activated by CNTs. Recently, potential mechanisms of CNT binding to PRRs have been suggested [34]. The impact of CNTs with regard to innate immune system activation is not currently understood. Such activation could lead to chronic inflammation and would be counterproductive to biomedical intervention in a clinical setting. Currently, there is a lack of data regarding the ability of CNTs to activate an innate immune response. | |
| To determine if CNTs activate an innate immune response and to identify mechanisms of CNT bioactivity, the mouse macrophagelike RAW264. 7 cells were used as a model of innate immune system activity in CNT exposure experiments. Increased extracellular levels of specific cytokines were observed upon MWCNT and exposure suggesting that the MWCNTs activate the innate immune system. Interestingly, the cytokine secretion profile was unique to MWCNTs compared to SWCNT and fullerene exposure suggestion a structuraldeterminant of activation. Additionally, MWCNT exposure resulted in the accumulation of the activated species of interferon regulatory factor 3 (IRF3) suggesting that this transcription factor is activated by MWCNT exposure. The extracellular concentration of the IRF3- induced cytokine IFN was significantly increased in response to MWCNT exposure suggesting that IRF3-modulated pathway is activated upon MWCNT exposure and may represent a specific mechanism of CNT bioactivity. The potential for innate immune system activation that may result in inflammation and compromised immunity must be determined to avoid unforeseen consequences of using CNTs for medical applications. | |
Results |
|
| Specific cytokines are secreted from RAW264.7 cells after MWCNT exposure | |
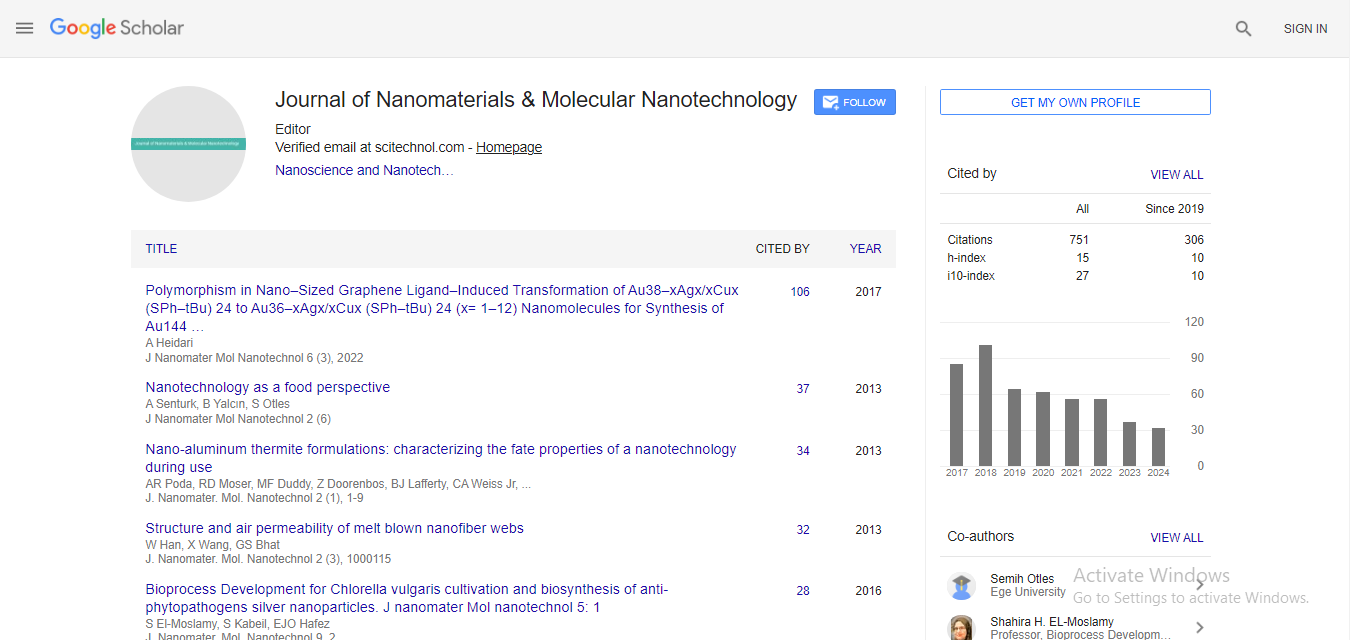
| To determine if an innate immune response is induced by exposure to CNTs, the mouse macrophage-like cell line, RAW264.7, was used for CNT exposure experiments. These cells are phagocytic and are known be involved in both the innate and cell-mediated immune response [35]. Macrophage cells represent a first line of defense against an immune system challenge and are typically activated by receptors that bind to pathogen associated molecular patterns (PAMPs). When activated, macrophage cells produce and secrete various cytokines that induce both intra- and extracellular signaling to facilitate the host immune response [36]. After exposure of RAW264.7 cells to MWCNTs, the extracellular levels of 16 cytokines were measured. MWCNT-1 exposure resulted in at least a 2-fold increase of 8 of the 14 chemokines/cytokines tested when compared to cells exposed to sonicated cell growth media alone (SM) (Figure 1A). The cytokines that were elevated upon exposure were Interleukin 1α (IL-1 α), IL- 2, IL-6, IL-9, interferon gamma (IFNγ), granulocyte macrophagecolony stimulating factor (GM-CSF), tumor necrosis factor alpha (TNF α, and regulated upon activation, normal T-cell expressed and secreted (RANTES). As a positive control for macrophage stimulation, cells were exposed to polyinosinic:polycytidylic acid (Poly I:C) or lipopolysaccaride (LPS) using a dose of 25 μg/mL and 100 ng/mL, respectively. Poly I:C and LPS are commonly used as PAMPs for activating an innate immune response. Exposure of RAW264.7 cells to Poly I:C or LPS increased the extracellular concentrations for all cytokines tested when compared to exposure to sonicated cell growth media (Figure 1B). Unlike Poly I:C and LPS exposure that increased the levels of all chemokines/cytokines tested, MWCNT-1 exposure increased the levels of only a subset of cytokines (Figure 1A). These data suggest that MWCNT-1 exposure results in the induction of a specific chemokines/cytokines that may result from the activation of specific pathways of the innate immune response. | |
| Figure 1: Relative extracellular cytokine levels after exposure of RAW264.7 cells to a sonicated cell growth media (SM), 50 µg/mL, or 500 µg/mL MWCNT suspensions in cell growth media b SM, 25 µg/ml polyinosinic:polycytidylic acid (Poly I:C), or 100 ng/mL lipopolysaccaride (LPS) in SM. Data are representative of three independent experiments and error bars represent standard error of the mean (SEM). | |
| All MWCNT preparations tested resulted in a similar chemokine/cytokine induction | |
| Profile: To determine if exposure to various MWCNT preparations also resulted in the induction of the observed cytokines from the initial experiments, RAW264.7 cells were treated with MWCNT-1, MWCNT-2, and MWCNT-3 suspensions. Prior to treatment with MWCNT and SWCNTs, the nanotube preparations were characterized for elemental composition and dimensions. The physical characteristics provided by the suppliers were confirmed for MWCNTs -2 and -3 by TEM analysis. However, MWCNT-1 preparation consisted of nanotubes that were greater in diameter than what was reported by the supplier (Figure 2). SWCNT were much larger in diameter than the 1-2nm reported by the supplier and might be due to aggregation. The chemical analysis for the various nanotube preparations was consistent with the analysis reported by the supplier. Various methods were employed for this characterization. After 24h treatment with MWCNT-1, MWCNT-2, and MWCNT-3 and SWCNTs cell culture supernatants were collected and the concentrations of extracellular cytokines were measured. Similar to MWCNT-1 treatment, MWCNT-2 and MWCNT-3 treatment resulted in increased levels of extracellular IL-6, IL-9, IFNγ, TNFα, and GM-CSF when compared to negative control. The observed TNFα induction upon MWCNT treatment is consistent with published results indicating that TNFα production is a consequence of MWCNT exposure [25,26,37]. RANTES levels were also increased upon exposure to all MWCNTs tested, however, this increase was much less than previously observed (Figure 3A). The induction was 2-fold for MWCNT-1 treatment, 2.2- fold for MWCNT-2 treatment and 1.5-fold for MWCNT-3 treatment. The induction of the extracellular levels of IL-1α 1.5-fold, 1.8-fold, and 1.9-fold for MWCNT-1 treatment, MWCNT-2 treatment, and MWCNT-3 treatment, respectively (data not shown) and was not consistent with previous MWCNT-1 treatment. Data represent cytokine levels that are normalized to the concentration observed after Poly I:C treatment to account for varying levels of RAW264.7 cell activation between experiments. These data indicated that regardless of supplier or size, MWCNT exposure induces production and secretions of a characteristic profile of cytokines. | |
| Figure 2: TEM images of nanotubes used in cell exposure experiments A)MWCNT-1, B) MWCNT-2, C) MWCNT-3, and D) SWCNT. | |
| Figure 3: Normalized fold induction of specific chemokines/cytokines in nanotube or fullerene treated cells. (A) RAW264.7 cells treated with sonicated cell growth media (SM) or 500µg/mL suspensions of MWCNT-1, MWCNT-2, or MWCNT-3 in cell growth media. (B) RAW264.7 cells treated with sonicated cell growth media (SM) or 500µg/mL suspensions of SWCNT or fullerene (C60) in cell growth media. Data are representative of three independent experiments and error bars represent standard error of the mean (SEM). | |
| To determine if the observed MWCNT-induced cytokine induction profile is unique to MWCNTs, RAW264.7 cells were treated with SWCNTs or C60 suspensions prepared by sonicating in cell growth media. After 24h treatment, extracellular chemokine/cytokine concentrations were determined using ELISA. Treatment with SWCNTs or C60 did not result in the induction of the same profile of cytokines observed after MWCNT treatment. SWCNT exposure increased the extracellular levels of 3 of the 8 cytokines (IL-6, IL-9, and IFNγ that were increased by MWCNT exposure. C60 exposure increased the extracellular level of 1 of the 8 cytokines (IL-9) that were increased by MWCNT exposure (Figure 3B). These data indicate that MWCNTs activate an innate immune response in RAW264.7 that results in an increase of a specific subset of cytokines. Furthermore, unlike the well-characterized PAMPs LPS and Poly I:C, MWCNT exposure did not induce global innate immune system activation suggesting that exposure results in the activation of a subset of signal transduction pathways in macrophage cells. | |
| MWCNT exposure results in the accumulation of phospho- IRF3 | |
| The increase in the levels of regulated upon activation, normal T-cell expressed, and secreted (RANTES), in response to MWCNT exposure is particularly interesting since transcription of the RANTES gene is known to be modulated by interferon regulatory factor 3 (IRF3) [38,39]. IRF3-mediated transcription is more specific than other transcription factors primarily by PAMP activation of the pathogen recognition receptors Toll-like Receptor 3 (TLR3) and TLR4 and is MyD88-independent [40-44]. We hypothesized that MWCNT exposure increases the levels of a specific subset of the cytokines tested by an IRF3-mediated mechanism. IRF3 is activated by C-terminal phosphorylation on serine396 mediated by TANK binding kinase 1 (TBK-1) and/or IKKi kinase [45]. To determine if activation of IRF3 represents a potential mechanism of the observed innate immune system activation by MWCNT exposure, the accumulation of IRF3 phosphorylated on the serine396 residue (phospho-IRF3) was determined in cells exposed to MWCNTs. MWCNT exposure resulted in the accumulation of phospho-IRF3 after a 3h treatment in a dosedependent manner suggesting that MWCNTs induce the activation of IRF3 (Figure 4A). These data suggest that MWCNTs exposure activates an IRF3-mediated pathway to increase the concentration of a subset of cytokines. | |
| Figure 4: (A) Interferon beta (IFNß) concentration in cell culture supernatants from RAW264.7 cells treated with sonicated cell growth media (SM), 25 µg/mL Poly I:C, 5 µg/mL, 50 µg/mL or 500 µg/mL MWCNT suspension in cell growth media. (B) Immunoblot analysis of interferon regulatory factor 3 (IRF3) protein expression and the accumulation of phosphorylated IRF3 (p-IRF3) in MWCNT treated RAW264.7 cells. Data are representative of three independent experiments and error bars represent standard error of the mean (SEM). *denotes statistically significant difference from SM treatment (P<0.05 calculated using student’s T-test). | |
| MWCNT exposure results in a dose-dependent increase in IFNβ levels | |
| A well characterized target for transcriptional activation by IRF3 is the type I Interferon, interferon beta (IFNβ) [38,39]. To provide further evidence that an IRF3-mediated pathway is activated in response to exposure, the concentration of IFNβ was determined in cell culture supernatants from cells exposed to varying concentrations of MWCNTs. A significant, dose-dependent increase in the extracellular concentration of IFNβ was observed upon MWCNT exposure (Figure 4B). Poly I:C is known inducer of IFNβ in RAW264.7 cells and was used as a positive control in this experiment. These data demonstrate that MWCNT treatment induced IRF3-mediated cytokine production and provide insight into specific innate immune system pathway activation by MWCNTs. Furthermore, these data provide evidence for the possibility that carbon-based nanomaterials engineered for biomedical applications harbor unique and specific bioactivity that must be fully understood prior to use. | |
Discussion |
|
| Carbon-based nanomaterials have unique properties that are ideal for many applications in biomedicine such as drug delivery and in the fabrication of medical devices [20]. However, the health consequences of introducing carbon-based nanomaterials into the human body for such purposes have not been adequately studied [11]. To gain a better understanding of the cellular and molecular consequences of exposure, we sought to determine if carbon-based nanomaterials activated signal transduction pathways that are characteristic of an innate immune response. | |
| In this contribution, we describe our use of the RAW264.7 mouse macrophage cell line to determine if carbon-based nanomaterial exposure initiates an innate immune response. Our data show that MWCNT exposure resulted in increased extracellular levels of specific cytokines and that this increase was unique to MWCNTs as C60 and SWCNTs did not increase the same subset of cytokines. Furthermore, MWCNT exposure resulted in the accumulation of activated IRF3 and the accumulation of IFN α, a known target of IRF3 mediated transcription. Taken together, these data suggest that pattern recognition receptors (PRRs) that are central to innate immune response may be activated by MWCNTs and mediate MWCNTinduced innate immune response via activation of specific signal transduction pathways. | |
| The great promise of using CNTs for biomedical applications will not be possible if such use results in a robust innate immune response. Such a response would be counterproductive in a clinical setting due to the induction of a chronic inflammatory state for the patient. Furthermore, a more complete understanding of the molecular mechanisms of innate immune system activation by CNTS much be achieved in order to mitigate these effects through CNT design. While our investigations identify the possibility of such a deleterious consequence of CNT exposure, it is important to note that much more experimentation is needed to determine if such exposures affect organisms in the same way. However, our results are consistent with others that show in various scenarios that CNTs elicit inflammation [46,47]. Furthermore, an understanding of the molecular mechanisms that underlie a CNT-induced innate immune response will facilitate the engineering and design of CNT to avoid such a response and therefore circumvent any deleterious consequences. | |
| The potential human health consequences associated with exposure to carbon nanotubes must be addressed before the promise of using these nanomaterials in everyday applications can be realized. The identification of a biomarker profile for MWCNT exposure will enable the development of a screening tool that can be used to assess the potential biological implications of carbon nanotube exposure. Furthermore, the activation of an IRF3-mediated innate immune system cell signaling pathway suggests that MWCNT may harbor bioactivity that results in unintended biological consequences associated with exposure. The unintended bioactivity of carbon-based nanomaterials must be investigated during product development and application. | |
Materials and Methods |
|
| Reagents | |
| Dulbecco’s Modified Eagle’s medium (DMEM), fetal bovine serum (FBS), (Gibco, Grand 242 Island, NY), IRF3 and phospho- IRF3 antibodies (Cell Signaling Technology, Beverly, MA), anti- rabbit IgG (Santa Cruz Biotechnology, Santa Cruz, CA), RAW264.7 mouse macrophage-like cells (ATCC, Manassas, VA), PolyI:C and LPS (Sigma-Aldrich, St. Louis, MO), MWCNT-1 (>95% MWCNTs, 20-30nm diameter and 5-7 μm length, BuckyUSA, Houston, TX), MWCNT-2 (>95-98% MWCNTs, 60-100 nm diameter and 5-15 μm length, cat # 1234NMG, NanoAmor, Los Alamos, NM) and MWCNT-3 (>95-98% MWCNTs, 10-30 nm diameter and 1-2 μm length, cat #1213NMGS, NanoAmor, Los Alamos, NM), SWCNT (90% SWCNTs, 1-2 nm diameter and 5-30 μm lenth, cat # 1284YJ, NanoAmor, Los Alamos, NM) | |
| Cell culture and treatments | |
| RAW264.7 cells were maintained in DMEM containing 10% heatinactivated fetal bovine serum supplemented with 100 IU penicillin/ ml and 10 g streptomycin/ml in 10 cm cell culture flasks at 37ºC, 5% CO2 and 95% humidity. Prior to treatment with carbon nanomaterials, RAW264.7 cells were grown overnight in 6 cm plates after plating at 1.5 × 106 cells/plate density. For all treatments, media was sonicated with the indicated dose of treatment at 40% power for ~ 2 minutes with shaking on ice to prevent localized heating. Immediately following sonication, suspended CNT treatments were added to cell cultures. | |
| ELISA | |
| RAW264.7 cells were exposed to 50 and 500 μg/mL suspensions of the above carbon nanomaterials for 24 hours. After 24h exposure, cell culture supernatants were collected and analyzed to determine the concentration of extracellular cytokines using the Q-Plex Mouse Cytokine Array (Quansys Biosciences, Logan, UT) following manufacturers protocol. Cell culture supernatants were cleared of carbon nanotube by spinning for 5 minutes spin at 10,000 X g in a microcentrifuge at 4°C and then stored at -80°C until the ELISA was performed. | |
| Immunoblotting | |
| Cells exposed to carbon nanomaterials for the indicated period of time were washed twice with cold PBS. After washing, whole cell protein extracts were obtained by physical removal of cells from plate with a cell scraper in the presence of radio-immunoprecipitation assay (RIPA) buffer (Pierce Biotechnology, Rockford, IL). Whole-cell protein extracts were obtained by 10,000 X g centrifugation maintained at 4ºC for 15 minutes. Protein conentation was determined using BCA assay (Pierce Biotechnology, Rockford, IL). 30 μg of total protein was resolved on 12% SDS-PAGE and transferred by electroblotting onto PVDF membrane. Membrane was blocked with 5% milk TBST (0.1% Tween) for 1hr at room temperature and then incubated overnight using a 1:10,000 dilution of anti-IRF3 or anti-phosho-IRF3 antibody at 4°C in 5% bovine serum albumin (Sigma-Aldrich, St. Louis, MO) in TBST. After incubation with primary antibody, membrane was washed 3 times for 5 minutes with TBST at room temperature. Secondary antibody (1:50,000) incubation was performed in 5% milk TBST at room temperature for 1hr followed by washing 3 times for 5 minutes in TBST. Antibody presence was detected using SuperSignal West Dura Extended Duration Substrate (Pierce Biotechnology, Rockford, IL) and visualized using a ChemiDoc 283 XRS (Bio-Rad, Hercules, CA). | |
Author’s Note |
|
| K.DH. and M.S.P. conceived and designed the experiments; K.D.H and N.H.M. performed the experiments; K.D.H and N.H.M analyzed the data; S.K.D. and M.S.P contributed reagents/materials/ analysis tools; K.D.H wrote the paper. | |
| Conflicts of Interest | |
| The authors declare no conflict of interest. Declaration of Conflicting Interests The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article. | |
Funding |
|
| This work was supported by Los Alamos National Laboratory LDRD-DR program and Agnew Postdoctoral Fellowship (N. H. Mack). | |
References |
|
|
|